Ibarra-Luna F. et al. RSCTyH, Vol. 1, Num.2, Julio 2023
Resumen: En la actualidad, las bacterias cada vez aumentan más su resistencia a los antibióticos, en el caso de las fluoroquinolonas estas inhiben la síntesis de ADN, esta inhibición se da por la interacción del antibiótico con el complejo formado por la unión del ADN a las dianas de las quinolonas, la ADN-girasa y la topoisomerasa IV. Este estudio reporta la presencia de los genes gyrA y gyrB en el genoma de la cepa tipo Gardnerella vaginalis UMB0386, estos genes proporcionan resistencia a las fluoroquinolonas, además, se analizó la relación evolutiva entre especies que presentan estos genes. Palabras clave: Gardnerella vaginalis, Filogenia, Fluoroquinolonas. |
Abstract: Currently, bacteria are becoming increasingly resistant to antibiotics, in the case of fluoroquinolones these inhibit DNA synthesis, this inhibition occurs by the interaction of the antibiotic with the complex formed by the binding of DNA to the targets of quinolones, DNA gyrase and topoisomerase IV. This study reports the presence of the gyrA and gyrB genes in the genome of the Gardnerella vaginalis type strain UMB0386, these genes provide resistance to fluoroquinolones, in addition, the evolutionary relationship between species that present these genes was analyzed. Keywords: Gardnerella vaginalis, Phylogeny, Fluoroquinolones. |
Recibido: 22/06/2023 Aceptado: 27/06/2023 Publicado: 29/07/2023
Introducción
Gardnerella vaginalis es un bacilo inmóvil, no encapsulado, puede presentar fimbrias, con una longitud de 0,5 a 1,5 µm, Gram negativo, anaerobio facultativo, catalasa y oxidasa negativo [1] .
La vaginosis bacteriana suele ser causada por esta bacteria, esto ocurre cuando se pierde el equilibrio entre la microbiota de la vagina, provocando que G. vaginalis crezca de manera exponencial. Gardner y Dukes reportaron esta bacteria aislada de mujeres con vaginitis inespecífica y fue identificada como Haemophilus sp. debido a su tinción Gram negativa y su nulo crecimiento en agar sin sangre [1].
Por otro lado, las fluoroquinolonas son agentes antimicrobianos relativamente recientes, inicialmente utilizadas contra bacterias Gram negativas, que posteriormente fueron modificadas químicamente para incrementar su espectro de acción sobre bacterias Gram positivas. La resistencia a las fluoroquinolonas es preocupante, debido a que las fluoroquinolonas son altamente eficaces para el tratamiento de algunas enfermedades graves producidas por bacterias multirresistentes en humanos [2].
En los últimos años aparecieron cepas resistentes a las fluoroquinolonas de tercera y cuarta generación, pero éstas siguen siendo eficaces para tratar esta infección. La aparición de resistencia a antimicrobianos es consecuencia natural del proceso evolutivo ante la mayor presión de selección por parte de la quimioterapia. La resistencia de los patógenos responsables de las ITS (infecciones de transmisión sexual) no se limita a los betalactámicos naturales, sino que incluye fármacos sintéticos como las fluoroquinolonas [3].
En el presente estudio se reportan genes relacionados a la resistencia a las fluoroquinolonas en el genoma de la cepa Gardnerella vaginalis UMB0386 asociada a la vaginosis bacteriana (VB).
Materiales y Métodos
Se descargó el genoma de la bacteria G. vaginalis UMB0386 de National Center for Biotechnology Information (NCBI). Posteriormente, se anotó en la plataforma de RAST (Rapid Annotations using Subsystems Technology) [4], para detectar los subsistemas y localizar los genes relacionados con resistencia a los antibióticos de G. vaginalis. Se descargaron y alinearon las secuencias de estos genes y se construyeron los árboles filogenéticos con MEGA [5], utilizando los métodos Neighbor-Joining y Maximum likelihood.
Resultados y Discusión
Anotación del genoma
El tamaño del genoma G. vaginalis UMB0386 fue 1.67 Mb (1,675,681 pb) con una cobertura genómica de 144x.
El genoma anotado presentó 355 (27%) secuencias asociadas a un subsistema y 981 (73%) secuencias no asociadas a un subsistema, por tal motivo las secuencias no lograron ser asignadas a un gen (figura 1). Se obtuvo de RAST la gráfica circular donde se observa la proporción que ocupa cada uno de los subsistemas del genoma de la cepa G. vaginalis UMB0386, el subsistema en el cual se pudo observar un mayor número de secuencias codificantes fue el de Carbohidratos (95), además, los subsistemas de latencia y esporulación, fagos, profagos, elementos transponibles y plásmidos presentaron solamente una secuencia codificante. En particular, el subsistema de virulencia, enfermedad y defensa contiene 14 secuencias codificantes, de las cuales 5 corresponden a resistencia a antibióticos y compuestos tóxicos.
Figura 1. Genes relacionados a subsistemas y su distribución en diferentes categorías en el genoma de la cepa G. vaginalis UMB0386
Obtención de los genes
Se identificaron dos genes de resistencia a antibióticos, en particular con resistencia a fluoroquinolonas (gyrA y gyrB).
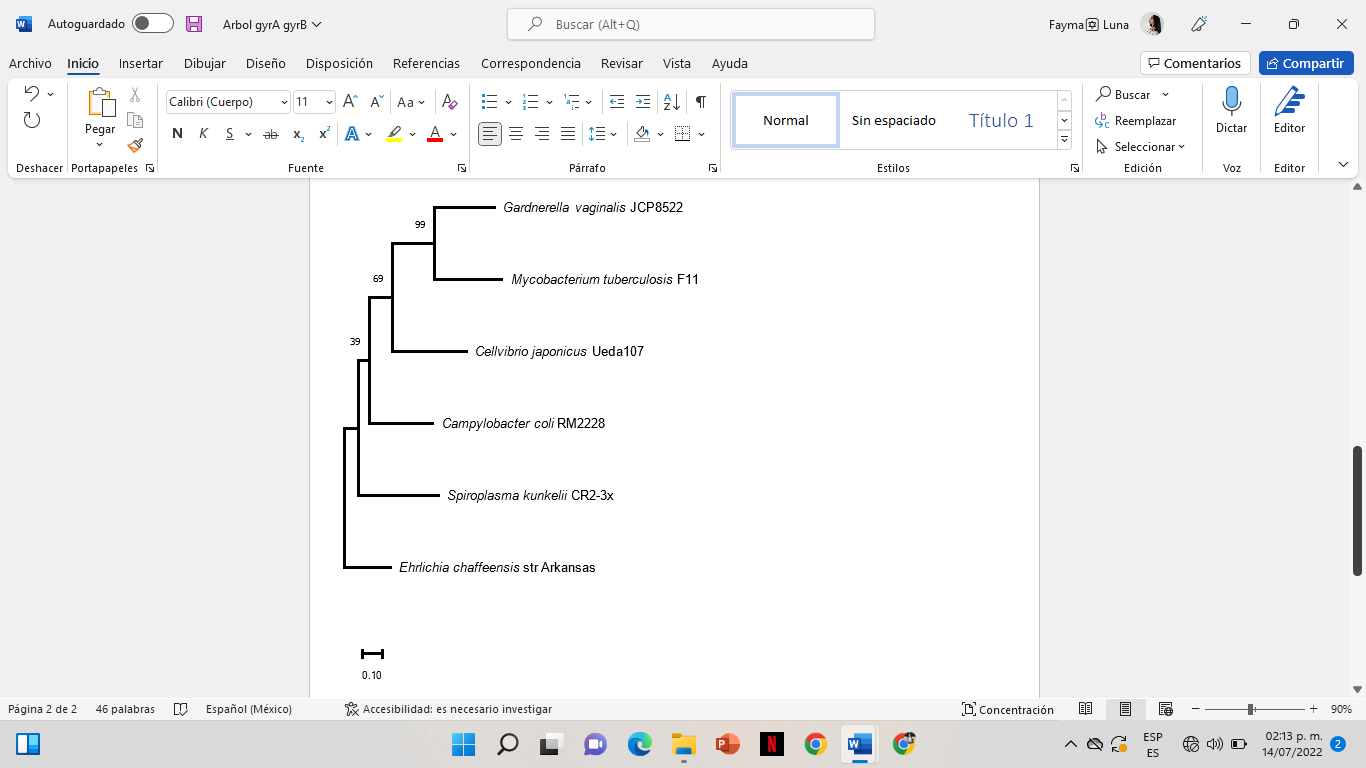
En la base de datos de RAST se comparó el genoma de la cepa de G. vaginalis UMB0386 y reveló el conjunto de genes con secuencias similares de los genes gyrA y gyrB en cinco especies distintas las cuales son Mycobacterium tuberculosis F11, Cellvibrio japonicus Ueda107, Campylobacter coli RM2228, Spiroplasma kunkelii CR2-3x y Ehrlichia chaffeensis str. Arkansas.
En 2005, se analizaron 17 aislamientos clínicos de Corynebacterium striatum y cepas de Corynebacterium amycolatum por el método E-test en placas de agar Mueller-Hinton. La cepa C. striatum ATCC 6940 se utilizó como cepa de control. Se caracterizaron las regiones determinantes de resistencia a quinolonas amplificadas de los genes gyrA de C. amycolatum y C. striatum. Se seleccionaron y analizaron cuatro mutantes de C. amycolatum resistentes a quinolonas in vitro [6].
En 2008, se evaluaron 348 aislamientos invasivos de S. aureus, 208 eran resistentes a meticilina y 140 sensibles a meticilina. Además, 155 aislamientos de S. aureus presentaron resistencia a la meticilina y todos los S. aureus sensibles a la meticilina. La confirmación de especie de estos aislamientos y la detección del gen de resistencia a meticilina (mecA) se realizó por ensayos de PCR múltiple. Las tasas de resistencia para ciprofloxacino, gatifloxacino y moxifloxacino en los 348 aislamientos de S. aureus fueron 55,4%, 54,9% y 52,6%, respectivamente. La alta resistencia a las fluoroquinolonas fue evidente en los aislamientos de S. aureus resistente a la meticilina, en los que las tasas de resistencia fueron del 92,3%, 91,8%, 91,3% y 87,5% para ciprofloxacino, levofloxacina, gatifloxacino y moxifloxacino, respectivamente [7].
Análisis filogenético
Figura 2. Árbol filogenético del gen gyrA con seis especies distintas, todas cuentan con resistencia a las fluoroquinolonas.
Se descargaron los genes gyrA y gyrB de cinco especies, los genes se alinearon con los genes de G. vaginalis UMB0386, posteriormente, se construyeron dos árboles filogenéticos con MEGA para comparar la distancia evolutiva entre las especies, en el árbol filogenético del gen gyrA se observa que su grupo externo es E. chaffeensis str. Arkansas y un clúster que presentó un 100% de Bootstrap, el árbol filogenético está compuesto por el clúster de G. vaginalis UMB0386, M. tuberculosis F11 y C. japonicus Ueda107, además de dos ramas independientes C. coli RM2228, S. kunkelii CR2-3x y el grupo externo E. chaffeensis str. Arkansas.
Figura 3. Árbol filogenético con genes gyrB, las 6 especies con resistencia a la fluoroquinolona.
En el segundo árbol filogenético (figura 3) se compararon genes de gyrB, se observó que su grupo externo es E. chaffeensis str Arkansas y de nuevo se observó un clúster y presentó coincidencia de los genes, en el primer clúster se obtuvo un 99% de coincidencia entre G. vaginalis UMB0386 y M. tuberculosis F11, se debe resaltar que la topología de los árboles es similar debido a que estos genes forman parte de un mismo operón.
En 2005, se seleccionaron y analizaron cuatro mutantes de C. amycolatum resistentes a quinolonas in vitro, cepas resistentes a quinolonas tanto in vivo como in vitro, las cuales mostraron altos niveles de resistencia a la fluoroquinolona en cepas con una doble mutación que conducía a un cambio de aminoácidos en las posiciones 87 y 91 o en las posiciones 87 y 88 de gyrA, mientras que las mismas mutaciones concomitantes en las posiciones de aminoácidos 87 y 91 en gyrA de C. striatum produjo altos niveles de resistencia a ciprofloxacino y levofloxacino, pero solo mostró un aumento moderado en la Concentración Mínima Inhibitoria (CIM) de moxifloxacino, lo que sugiere que otros mecanismos de resistencia a quinolonas podrían estar involucrados en la resistencia a moxifloxacino [6].
Conclusión
Se detectaron los genes gyrA y gyrB en el genoma de G. vaginalis UMB0386, los cuales les proporcionan resistencia a antibióticos, especialmente a las fluoroquinolonas. Este grupo de antibióticos formado por ciprofloxacino, levofloxacino, moxifloxacino, norfloxacino y ofloxacino, tienen efecto bactericida, mediante la inhibición de síntesis de ADN bacteriano. Se encontró que no solo G. vaginalis UMB0386 tiene este efecto de resistencia si no también se puede encontrar en cepas de C. japonicus Ueda107, C. coli RM2228, S. kunkelii CR2-3x, E. chaffeensis str. Arkansas y M. tuberculosis F11, esta última presentó una estrecha relación filogenética con G. vaginalis en los árboles filogenéticos que presentaron una topología similar. Finalmente, la detección de genes de resistencia a las fluoroquinolonas en el genoma de G. vaginalis UMB0386 es fundamental para comprender y abordar el problema creciente de la resistencia antimicrobiana. La capacidad de detectar y monitorear estos genes es crucial para la implementación de estrategias de control y prevención eficaces.
Referencias
[1] Hernández, F. (1998). Gardnerella vaginalis Mobiluncus en la etiología de la vaginosis bacteriana. Revista Costarricense de Ciencias Médicas, 19(1-2), 107-116. Retrieved June 13, 2022, from http://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S0253-29481998000100010&lng=en&tlng=es.
[2] Briseño-Torres, L, Narváez-Bravo, C, Rodas-González, A, Wittum, T, & Hoet, A. (2007). Resistencia a las Fluoroquinolonas y Otros Antimicrobianos en Cepas de Salmonella spp. Aisladas en el Procesamiento de Pollo Entero.. Revista Científica, 17(5), 521-528. Recuperado en 15 de agosto de 2022, de http://ve.scielo.org/scielo.php?script=sci_arttext&pid=S0798-22592007000500013&lng=es&tlng=es
[3] Shaskolskiy, B, Dementieva, E, & Gryadunov, D. (2016). Resistencia farmacológica en bacterias asociadas con infecciones de transmisión sexual y vaginosis. Laboratorios Bagó. Recuperado 16 de julio de 2022, de https://www.bago.com.ar/vademecum/bibliografia/resistencia-farmacologica-en-bacterias-asociadas-con-infecciones-de-transmision-sexual-y-vaginosis/
[4] Aziz, RK, Bartels, D, Best, AA, Matthew, D, Disz, T, Edwards, RA, Formsma, K, Gerdes, S, Glass, E, Kubal, M, Meyer, F, Olsen, GJ, Olson, R, Osterman, AL, Overbeek, RA, McNeil, LK, Paarmann, D, Paczian, T, Parrello, B, Pusch, GD, Reich, C, Stevens, R, Vassieva, O, Vonstein, V, Wilke, A, Zagnitko, O. (2008). The RAST Server: Rapid Annotations using Subsystems Technology. 26/6/2022, de BMC Genomics Sitio web: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2265698
[5] Tamura, K, Stecher, G, Kumur, S. (2021). MEGA11:
Molecular Evolutionary Genetics Analysis version 11. 27/6/2022, de Molecular Biology and Evolution Sitio web: https://academic.oup.com/mbe/article/38/7/3022/6248099?login=false
[6] Sierra, M, Martínez, L, Vázquez, F, Giralt, E & Vila, J. (2005). Just a moment. . . ASM Journals. Recuperado 15 de julio de 2022, de https://journals.asm.org/doi/10.1128/AAC.49.5.1714-1719.2005
[7] Hidalgo, M, Reyes, J, Cárdenas, A, Díaz, L, Rincón, S, Vanegas, N, Díaz, P, Castañeda, E, & Arias, C. (2008). Perfiles de resistencia a fluoroquinolonas en aislamientos clínicos de cocos Gram positivos provenientes de hospitales colombianos, 1994-2004. Biomédica, 28(2), 245-251. Retrieved July 15, 2022, from http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-41572008000200013&lng=en&tlng=es